You are on the first web site of Biotic Regulation. Since February 14, 2008 it is no longer updated. Welcome to www.bioticregulation.ru! |
Empirical evidence for the biotic regulation concept
Climate instability |
|
| next section | |
Independent studies [1] suggest that the mean global temperature of the Earth's surface has remained within the life-compatible interval from +5 to +25° C during the whole period of life existence, i.e. for the last 3,85 billion years. Following Charles Darwin, the modern biological theory states that species adapt genetically to changing environments. Such an approach to species-environment interaction takes the existence of life-compatible environmental conditions on the planet for granted. It implicitly assumes that either
- Environmental conditions fluctuate within a life-compatible corridor, where they are kept by some stabilising physical factors. For example, the life-compatible range of surface temperatures may be dictated by the position that our planet occupies in the solar system.
- The fact that during the last 3,85 billion years the environmental conditions have not fallen out of the life-compatible range owes itself completely to a lucky chance.
Such a consideration is absolutely essential for the modern biological theory. If it appeared that the Earth's environment, left to itself, rapidly degraded to a life-incompatible state under the influence of abiotic factors, then the very notion of life must include environmental control. The ability of environmental regulation must be as indispensable in the definition of living systems as are now the abilities to reproduce, to feed, to form populations. These abilities themselves should then be viewed as various manifestations of the major property of living systems - environmental regulation. Indeed, in order to regulate the environment, a living system must have a metabolism (i.e. it has to feed). In order to sustain the regulation over longer periods of times a living system has to be able to make new copies of itself instead of old ones (i.e. to reproduce). In order to prevent the information needed for environmental regulation from decay, living systems must form populations and force out individuals with degraded program of regulation, etc.
In other words, the nature of stability of life-friendly environmental characteristics is an issue directly related to the theoretical consideration of living systems in modern biology.
Contrary to the above implicitly accepted propositions, analysis of the nature of climate stability on Earth [2] indicates that the modern climate of Earth with its predominantly liquid hydrosphere is physically unstable. It means that it is impossible to find a lifeless planet with the same external physical characteristics as on Earth, where an ocean similar to the terrestrial one would be existing for a geological period of time. Governed by physical regularities, such a planet would rapidly evolve to a state of either complete evaporation of the hydrosphere (with a catastrophic greenhouse effect) or to a state of complete glaciation of the planet's surface (with a negligible greenhouse effect and an unacceptably high value of albedo).
Both these states are unfit for life. The temperature of the planet's surface is a function of three variables, solar constant (i.e. how much energy comes from the Sun per unit surface), greenhouse effect and albedo. Greenhouse effect describes how much of thermal radiation emitted from the planet's surface is effectively returned to the surface by greenhouse substances in the atmosphere. The greenhouse effect leads to additional heating of the planet's surface. For example, on Earth without the greenhouse effect the global mean surface temperature would have been 33°C lower than it is now and reached -18°C [3]. Albedo represents the fraction of the incoming solar radiation that is reflected by the planet back into space. If albedo is 100%, the planet does not receive energy at all. The modern value of albedo on Earth is 30%.
The value of solar constant is determined by the position of the planet in the solar system. The further from the Sun, the less energy that the planet receives. However, by varying the values of albedo and greenhouse effect that are completely determined by the inherent environmental characteristics of the planet, it is possible to maintain an almost arbitrary value of the surface temperature at any planet. For example, to keep warm a planet very distant from the Sun, one needs to add large amounts of greenhouse substances to the planet's atmosphere. On the contrary, in order to cool a planet which is too close to the Sun, it is necessary to increase its albedo. Finally, on a planet which seems to be at a favourable distance from the Sun, alterations of environmental characteristics and the related changes in the greenhouse effect and albedo may yield a life-incompatible value of surface temperature.
On Earth the major greenhouse gas is the water vapour [3]. The concentration of water vapour in the atmosphere depends on surface temperature in accordance with Clausius-Clapeyron equation [4]. With growing temperature the concentration of water vapour grows rapidly, because more water evaporates from the ocean and rests in the atmosphere. As a result, the greenhouse effect grows, which works to increase the surface temperature further and so on. Similarly, an occasional drop of the surface temperature leads to depletion of water vapour from the atmosphere, decrease of the greenhouse effect, further drop of the surface temperature and so forth.
The analysis of the rates of these positive destabilising feedbacks [2] show that a climate with a liquid hydrosphere is unstable with respect to spontaneous transition to either complete evaporation of all the planet's water (with surface temperatures higher than +400°C like on Venus) or complete glaciation of the planet (with temperatures lower than -80°C and a negligible greenhouse effect as on Mars). Moreover, these changes are likely to be nearly instantaneous on a geological scale, covering time periods of about thousand years.
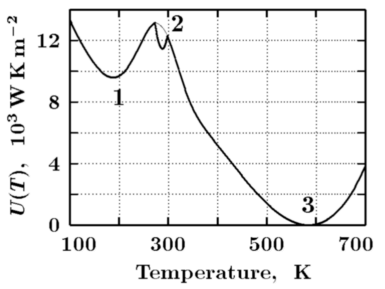
In the absence of physical mechanisms that could account for the maintenance of a life-compatible temperature during the last four billion years, one has to accept that the observed stability of the climate with liquid hydrosphere is of biotic nature. Life appears therefore to be able to control the values of albedo and greenhouse effect compensating for their fluctuations and buffering the unfavourable temperature changes. Such a situation corresponds to the existence of a stable biotic pit in the vicinity of the modern global mean surface temperature. A model example of such a pit is shown in Fig. 1, state 2, thick line.
Anthropogenic cultivation of natural ecosystems is equivalent to shallowing of the biotic pit followed by its ultimate disappearance. This may end in a transition of the Earth's climate to either of the two stable but life incompatible states depicted in Fig. 1. Such a transition would correspond to rapid and irreversible sliding down the potential hill to either of the two life-incompatible pits.
Species discreteness |
to the top |
| previous section | |
| next section |
There is another important line of evidence in favour of the biotic regulation concept. The available paleodata testify for morphologic and, consequently, genetic [5] constancy of all species-specific characteristics during the whole period of the existence of most of the species studied [6]. In most cases, a species arises in the paleorecord as a narrow set of similar morphological forms, persists like this during the whole period of the species' existence (several million years on average) and disappears. A new related species that occupies its place in the record is separated from the old one by a considerable gap in the total set of morphological characteristics. In other words, no transient forms that would viaduct two discrete acts of speciation are ever observed.
The extant species also demonstrate strict discreteness. There are neither intermediate forms between related species, nor processes of their formation being observed. Hybrid zones that are sometimes considered as possible seedlings of evolutionary process, are strongly restricted and occupy a few per cent of the biosphere at maximum [7].
Evidence for discreteness of species remains a topic largely discussed by paleobiologists, with little consideration by theoretical biologists. This is partly because of the fact that the observed patterns cannot be consistently explained if one assumes that species exist in the process of adaptation to changing environmental conditions. Why, in spite of the assumed continuous process of adaptation to an ever-changing environment, does the majority of all species retain morphological discreteness both in space and time?
The biotic regulation concept allows to explain this pattern. If one accepts that the biological species organised into ecological communities control their environment, it becomes clear that the process of biotic regulation demands large amounts of information about how to stabilise the environment and what environment should be stabilised. It means that the genetic information of species should include the information about characteristics of the optimal environment. Definitely, for the biotic regulation to be possible, the genetic information of species may not change in response to environmental fluctuations. Instead, this information should ensure a stabilising response of the species inside the community which restores the environment back to the optimal state. This attitude unambiguously predicts that the species should remain unchanged - genetically and, consequently, morphologically - during most part of their existence.
The revealed tempo and mode of evolutionary processes support the view that the evolution of species is not a result of gradual accumulation of relatively frequent minor modifications of the hereditary program of a species. Instead, the evolutionary process is discontinuous and is due to infrequent but radical quantum changes of the discrete genetic programs of species.
The continuous "daily" process of mutations and other hereditary genetic changes that supports intraspecific genetic polymorphism bears therefore no relation to the evolutionary process. Rather, it can be conceived as random deviations from the stable normal hereditary program of the species. Random genetic deviations accumulate due to the mutational process and erase the genetic information of the species. The number of such deviations cannot increase infinitely, but is limited by the stabilising selection [8]. Individuals with too many genetic deviations are forced out from the population. Thus, under natural conditions the genetic program of species, necessary for biotic regulation, is prevented from decaying. This fact explains the observed constancy of species-specific characteristics in time.
More |
to the top |
| previous section | |
| next section |
We have briefly outlined the two major domains of empirical evidence, climate instability and species discreteness, that form the logical basis for the biotic regulation concept. However, there are many other lines of evidence pointing in the same direction. If you are interested, we suggest that you read the first chapter from Gorshkov et al. (2000) Biotic regulation of the environment: Key issue of global change, where the general overview of the concept is given, see in particular Section 1.8. Read also the publications on the topic available here. Finally, there is a somewhat more detailed discussion of how the biotic regulation concept relates to the major paradigms of the modern biology, in particular, to the concepts of genetic adaptation and nutrient limitation, see here.
If you have any comments, we welcome you to share your thoughts in our on-line discussion area.
References |
to the top |
| previous section | |
[1]
Savin S. (1977) The history of the Earth's surface temperature during the past 160 million years. Ann.Rev. Earth Planetary Sci., 5, 319-355.
Watts J.A. (1982) The carbon dioxide question: Data sampler. In: Clark W.C. (Ed.) Carbon Dioxide Review. Clarendon Press, New York.
Berggren W.A. and Van Couvering J.A. (Eds.) (1984) Catastrophes and Earth History: The New Uniformitarianism. Princeton University, New York.
back to the text
[2]
Gorshkov V.G., Makarieva A.M. (2002)
Greenhouse effect dependence on atmospheric
concentrations of greenhouse substances and the nature of
climate stability on Earth. Atmos. Chem. Phys. Discuss.,
2, 289-337.
Abstract, PDF and discussion,
Gorshkov V.G., Gorshkov V.V., Makarieva A.M. (2000) Biotic regulation of the environment: Key issue of global change. Springer, London, 367 pp.
Contents,
Chapter 1.
Makar'eva A.M., Gorshkov V.G. (2001) The greenhouse effect and the
stability of the global mean surface temperature.
Doklady Earth Sciences, 377(2), 210-214.
Full text (PDF, 173 Kb).
Makarieva A.M. (2000) Biotically maintained stability of the Earth's mean global surface temperature. Petersburg Nuclear Physics Institute, Preprint No. 2384, 42 pp.
Full text (PDF, 0.4 Mb).
Gorshkov V.G., Makarieva A.M. (2000) Environmental safety,
climate stability and the non-perturbed biota.
Global Change NewsLetter, 43, 24-25.
Full text (PDF, 0.1 Mb).
Gorshkov V.G., Makarieva A.M. (2001) Diffusion of thermal photons
in the atmosphere, pp. 141-142 in: Dakhno L.G. (Ed.) PNPI XXX
(Petersburg Nuclear Physics Institute XXX-th Anniversary).
Scientific Highlights.
Theoretical Physics Division. PNPI, Gatchina, 145 pp.,
ISBN 5-86763-043-9.
Full text (PDF, 0.1 Mb).
back to the text
[3]
Mitchell J. (1989) The "greenhouse" effect and climate change.
Rev. Geophys., 27, 115-139.
back to the text
[4]
Raval A., Ramanathan V. (1989) Observational determination of
the greenhouse effect. Nature, 342, 758-761.
back to the text
[5]
Jackson J.B.C. (1990) Evolutionary significance of
morphospecies: a test with cheilostome Bryozoa. Science,
248, 579-583.
back to the text
[6]
Stanley S.M. (1979) Macroevolution: pattern and process. San
Francisco: Freeman.
back to the text
[7]
Raven P.H., Johnson G.B. (1988) Understanding Biology. Times
Mirror, Mosby Coll. Pub., St. Louis.
back to the text
[8]
Gorshkov V.G., Makar'eva A.M. (1997) Dependence of heterozygosity on
body weight in mammals. Doklady Biological Sciences, 355, 384-386.
Full text (PDF, 127 Kb).
Gorshkov V.G., Makar'eva A.M. (1999) Haldane's Rule and somatic mutations.
Russian Journal of Genetics, 35(6), 611-617.
Abstract,
full text (PDF, 184 Kb).
Makarieva A.M. (2001) Variance of protein heterozygosity in different
species of mammals with respect to the number of loci studied.
Heredity, 87(1), 41-51.
Abstract,
full text (PDF, 0.4 Mb),
the dataset (HTML, 0.1 Mb).
back to the text
|